
O. Semyachkina-Glushkovskaya, A. Abdurashitov, A. Pavlov, A. Shirokov, N. Navolokin, O. Pavlova, A. Gekalyuk, M. Ulanova, N. Shushunova, A. Bodrova, E. Saranceva, A. Khorovodov, I. Agranovich, V. Fedorova, M. Sagatova, A. E. Shareef, C. Zhang, D. Zhu, V. Tuchin, "Laser speckle imaging and wavelet analysis of cerebral blood flow associated with the opening of the blood–brain barrier by sound," Chin. Opt. Lett. 15, 090002 (2017)
- Chinese Optics Letters
- Vol. 15, Issue 9, 090002 (2017)
Abstract
The blood–brain barrier (BBB) is a highly selective gatekeeper, which controls the passage of blood-borne agents into the brain tissues protecting the brain against pathogens. In the early 1940s in experiments
Solute clearance across the BBB is an important function of cerebral blood flow (CBF). The rate of blood flow within the brain parenchyma, where the BBB is located, is 130-fold greater than the rate of blood flow within the choroid plexus, where the leaky choroid barrier is resented[
The cerebral circulation keeps a stable and unique extracellular environment within the neurovascular unit, which is essential for the BBB function. Clinical and experimental data suggest that regulation of the BBB is often impaired in pathological brain conditions associated with diverse vascular abnormalities, such as ischemia, stroke, brain tumors, or trauma leading directly to the malfunction of the neurovascular unit and long-lasting changes in neuronal activity[
Sign up for Chinese Optics Letters TOC. Get the latest issue of Chinese Optics Letters delivered right to you!Sign up now
Despite accumulating evidence for the crucial role of CBF in the BBB function, there is limited information about the changes of CBF related to the BBB disruption. Here, we studied changes of CBF in mice associated with the opening BBB by sound, using a proposed algorithm for laser speckle contrast imaging (LSCI) and wavelet analysis of the venous and microvascular components of the CBF.
Experiments were carried out in adult male mice that were two months old, using four groups: (1) no sound, the control group, (2) 90 min, (3) 4 h, and (4) 24 h after sound-stress. Each group included 10 mice. All procedures were performed in accordance with the “Guide for the Care and Use of Laboratory Animals”[
To induce the opening of BBB, we used audible sound (110 dB, 370 Hz): 60 s sound, then 60 s pause, and this cycle repeated during 2 h. This procedure was performed using a sound transducer in a Plexiglas® chamber (the volume
A polyethylene catheter (PE-10 tip, Scientific Commodities INC., Lake Havasu City, Arizona) was inserted into the right femoral vein for Evan’s Blue (EB) dye (Sigma Chemical Co., St. Louis, MI, USA) intravenous injection in a single bolus dose (2 mg/25 g mouse, 1% solution in physiological 0.9% saline). The EB circulated in the blood for 30 min. The implantation of the catheter was performed under the inhalation anesthesia (2% isoflurane, 70%
Fluorescein rhodamine (FITC)-dextran 70 kDa was used as the additional method for characterization of the BBB permeability to high-weight molecules. FITC-dextran was injected intravenously (4 mg/25 g mouse, 0.5% solution in 0.9% physiological saline, Sigma) and circulated 2 min. Afterwards, the mice were killed by decapitation, and the brains were quickly removed, fixed in 4% paraformaldehyde (PFA) for 24 h, cut into 50 μm thick slices on a vibratome (Leica VT 1000S Microsystem, Germany), and analyzed on a confocal microscope (Nikon TE 2000 Eclipse, Tokyo, Japan).
All mice were decapitated after the performed experiments. The samples of the brain were fixed in 10% buffered neutral formalin. The formalin fixed specimens were embedded in paraffin, sectioned (4 μm), and stained with haematoxylin and eosin. The histological sections were evaluated by light microscopy (mVizo-103, Russia).
A home-made system for the LSCI was used for measurements of the perfusion of brain tissues. The raw speckle images were recorded under the following conditions: light source—HeNe laser with the wavelength 632.8 nm; image sensor—CMOS camera Basler acA2500-14 gm; imaging lens—Computer M16140-MP2 16 mm at F-number equal to six that corresponds to speckle/pixel size ratio of around two; exposure time
![]()
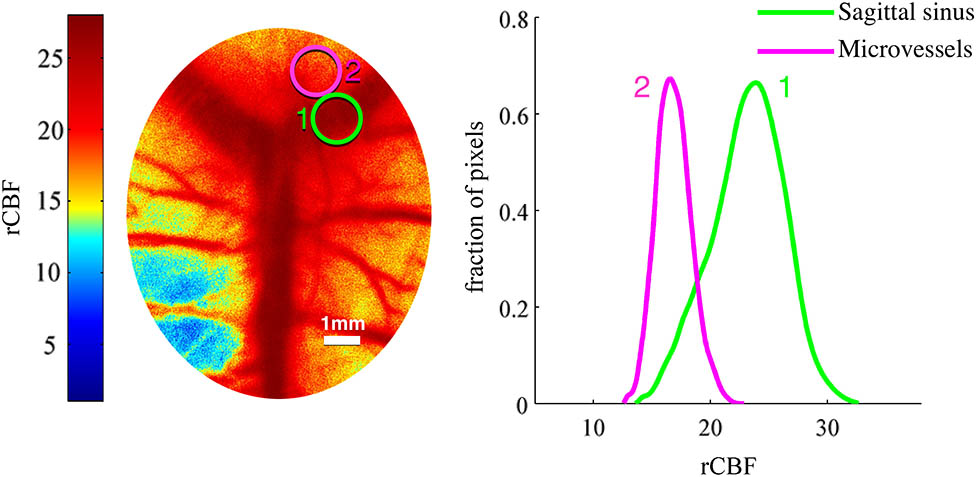
Figure 1.(Color online) (A) Two ROIs are overlaid onto LSCI image and (B) normalized histograms of these ROIs.
To characterize changes of the CBF, we used the wavelet-based multifractal analysis[
All skeleton lines are extracted by a selection of local maxima and minima of the wavelet-transform coefficients
The position of the singularity spectrum
Sign up for Chinese Optics Letters TOC. Get the latest issue of Chinese Optics Letters delivered right to you!Sign up now
The results were reported as
We studied the effect of sound on the BBB permeability using classical methods: spectrofluorometric and confocal assay of extravasation of high-weight molecules, such as EB 68.5 kDa (bound to serum albumin) and dextran 70 kDa, respectively. Both of these tracers do not permeate the intact BBB, and, therefore, they are widely used for the evaluation of the BBB disruption[
The EB concentration in the brain of intact mice (without sound) was about zero (
![]()
Figure 2.Spectrofluorometric assay of EB extravasation from the blood into the brain parenchyma suggesting the increase in the BBB permeability to EB in 90 min after sound with afterward-rapid normalization of BBB leakage in 4 h. Asterisks indicate significant changes (
For dextran 70 kDa, it was clearly shown that in the normal state (before sound), the BBB was not permeable for this tracer [Fig.
![]()
Figure 3.(Color online) Confocal imaging of BBB permeability to dextran 70 kDa in mice before and after sound: (A) no extravasation of dextran 70 kDa before sound; (B) and (C) extravasation of dextran 70 kDa in 90 min and 24 h after sound, respectively [defined as red clouds around the sagittal sinus, which is a main cerebral vein (B)], and a group of microvessels, including venules, draining the blood into the cerebral vein (C).
It is important to note that during all of the time of observation when the BBB is opened (90 min and 24 h after sound), we discovered the high permeability of cerebral vessels to dextran in both venous and microvascular types of vasculature. Figure
Figure
![]()
Figure 4.Histological analysis of BBB permeability to solutes of small molecular weight: (A) before sound, no solute extravasation; (B) in 90 min after sound, the vasogenic edema observed suggests the high BBB leakage for the water and other solutes with their accumulation in space between cerebral microvessels and the brain parenchyma; (C) in 24 h after sound-stress, the vasogenic edema is still observed.
The statement that the anatomic site of the BBB is cerebral microvessels is based predominantly on the data of electronic microscopy and immunohistochemistry obtained in 40 years of the last century, i.e., in
Figure
![]()
Figure 5.(Color online) LSCI for monitoring of the CBF of the cerebral veins and microvessels in four groups: control–(without sound); 90 min after sound—the reduction of venous CBF associated with the opening BBB; 4 h after sound—normalization of the venous CBF with recovery of the BBB; 24 h after sound—the repeated decrease in the venous CBF associated again with the opening BBB. No significant changes in the CBF on the level of microcirculation in different times after sound.
There were no pronounced and statistically significant changes in the CBF on the level of microvessels, probably due to compensatory mechanisms via distribution of blood in the cerebral veins as capacity link of the CBF.
In the previous work, the effectiveness of the wavelet analysis of CBF to discriminate the sensitivity of different vasculature components to stress during the development of vascular catastrophes in the brain was demonstrated[
The results of this series of experiments showed similar data to the LSCI: the BBB opening in 90 min and 24 h after sound-stress was associated with stronger changes of the mean Hölder exponent H quantifying CBF data in the cerebral veins compared to much weaker changes in the microvessels (Fig.
![]()
Figure 6.(Color online) Wavelet-based analysis of the changes of the CBF in the cerebral veins (orange) and microvessels (violet) before and after sound: the control (without sound); in 90 min and 24 h elapsed after sound-stress, the disruption of BBB function is seen; in 4 h, the recovery of the BBB function is seen. Asterisk indicates significant changes (
For CBF measurements performed in 4 and 24 h, the observed changes are less essential at both the macroscopic level of cerebral veins (
The revealed changes in the mean Hölder exponent have a relation with the corresponding changes in the decay of the correlation function and the power-law behavior of the spectral density. The latter characteristics, however, are difficult to estimate directly based on short and nonstationary data series acquired in physiological experiments. Due to this, the application of the wavelet-based multifractal formalism provides an indirect way of quantifying spectral and correlation properties of CBF, using a quite small amount of data.
The application of LSCI as a potential useful tool for the analysis of the BBB function is discussed in the review of Pandey
It should be noted that confocal imaging of the BBB disruption shows the high permeability of microvessels, including large (the sagittal sinus) and small cerebral veins [Figs.
However, now it is difficult to make a comparison with the other works because there is no reliable data for the CBF behavior associated with the BBB disruption due to the lack of research in this area. One group of authors, by using laser Doppler flowmetry, showed different changes in the CBF associated with the different sources for the transient BBB opening: the bile salt deoxycholic acid-induced BBB injuries are accompanied by an increase in the arterial and venous components of the CBF, while trombotic-induced BBB disruption is associated with an increase in the arterial but a decrease in venous cerebral circulation[
The data presented and discussed clearly show that more detailed studies of CBF related to the BBB disruption may shed light on a better understanding of the role of cerebral circulation in the regulation of environment within the neurovascular unit and for the BBB function. This new knowledge will contribute to the development of a new strategy for a therapeutic window of correction of the BBB disruption in pathological brain conditions associated with vascular abnormalities leading, to the malfunction of the neurovascular unit and long-lasting changes in neuronal activity.
References
[1] T. Broman. Acta Psych. Et Neurol., 16, 1(1941).
[2] U. Friedemann. Physiol. Rev., 22, 125(1942).
[3] W. M. Pardridge. Expert Opin. Drug Delivery, 13, 963(2016).
[4] R. Spector, R. F. Keep, S. R. Snodgrass. Exp. Neurol., 267, 78(2015).
[6] O. Tomkins, O. Friedman, S. Ivens, C. Reiffurth, S. Major. Neurobiol. Dis, 25, 367(2007).
[7] N. J. Abbott, A. Friedman. Epilepsia, 53, 1(2012).
[8] E. A. Neuwelt. Neurosurgery, 54, 131(2004).
[9] D. Fernandez-Lopez, J. Faustino, R. Daneman, L. Zhou, S. Y. Lee. J. Neurosci., 32, 9588(2012).
[10] . Guide for the care and use of laboratory animals.(2011).
[12] A. K. Dunn. Ann. Biomed. Eng., 40, 367(2012).
[13] J. F. Muzy, E. Bacry, A. Arneodo. Phys. Rev. Lett., 67, 3515(1991).
[14] J. F. Muzy, E. Bacry, A. Arneodo. Int. J. Bifurcation Chaos., 4, 245(1994).
[15] A. N. Pavlov, O. N. Pavlova. Tech. Phys. Lett., 34, 306(2008).
[16] P. Ch. Ivanov, L. A. Nunes Amaral, A. L. Goldberger, S. Havlin. Nature., 399, 461(1999).
[17] A. Hoffmann. Transl. Stroke Res., 2, 106(2011).
[18] S. Nag. Meth. Mol. Med., 89, 133(2003).
[21] P. K. Pandey, A. K. Sharma, U. Gupta. Tissue Barriers, 4, e1129476(2016).
[25] M. Hedtjärn, A. L. Leverin, K. Eriksson, K. Blomgren, C. Mallard, H. Hagberg. J. Neurosci., 22, 5910(2002).


Set citation alerts for the article
Please enter your email address

© Copyright 2018-2021 | Chinese Laser Press. All Rights Reserved 沪ICP备15018463号-20