
Jianyong Wang, Junchao Fan, Bo Zhou, Xiaoshuai Huang, Liangyi Chen, "Hybrid reconstruction of the physical model with the deep learning that improves structured illumination microscopy," Adv. Photon. Nexus 2, 016012 (2023)
- Advanced Photonics Nexus
- Vol. 2, Issue 1, 016012 (2023)
Abstract
1 Introduction
Superresolution (SR) fluorescence microscopy provides nanoscale resolution for studying subcellular structures and biological processes.1
Various physical model-based restoration methods have been developed to suppress SIM artifacts, such as total variation regularization,19 notch filtering,16,18 high-fidelity (HiFi)-SIM,20 and joint space and frequency reconstruction-SIM.21 Using spatiotemporal continuity as the prior knowledge, we have developed an iterative restoration method based on the Hessian regularization term (Hessian-SIM) that suppresses artifacts due to the amplification of random noise.18 However, other artifacts persist, such as hammer-stroke and honeycomb artifacts due to the out-of-focus background17,18,20 and artifacts due to illumination scattering,22 which cannot be suppressed completely by model-based methods. Deep neural networks can approximate arbitrary functions with infinitesimal errors to extract high-dimensional features from low-resolution and low-quality images.23 Therefore, researchers have proposed end-to-end deep-learning (DL)-based reconstruction algorithms to suppress different artifacts indiscriminately with low SNR raw SIM images.24
To combine the advantages of both methods, we try to balance the reconstruction fidelity of traditional methods and the artifacts suppression of DL methods. However, suppose these two parts are combined into one objective function to achieve simultaneous optimization; in that case, the network is required to calculate the partial differential28 of the input images rather than the normal network weights. Therefore, we utilized the total deep variation (TDV) network as a regularizer in the reconstruction objective function. By combining the physical SIM reconstruction procedure with the TDV regularizer,28 we propose a hybrid restoration method (TDV-SIM) to suppress artifacts and maintain resolution simultaneously. On processing images of different cellular structures, TDV-SIM retains the actual signals better than the pure DL methods while removing artifacts more effectively than the model-based methods.
Sign up for Advanced Photonics Nexus TOC. Get the latest issue of Advanced Photonics Nexus delivered right to you!Sign up now
2 Methods
2.1 Principle and Parameter Selection of TDV-SIM
For SIM imaging, the sample is excited by sinusoidal illuminations with different pattern orientations and phases. The raw images contain low- and high-frequency information, which need to be separated and reassembled in SIM reconstruction.16,18,19 We transformed the SIM reconstruction into an optimization problem and constructed an objective function [Eq. (1)] composed of the fidelity term
![]()
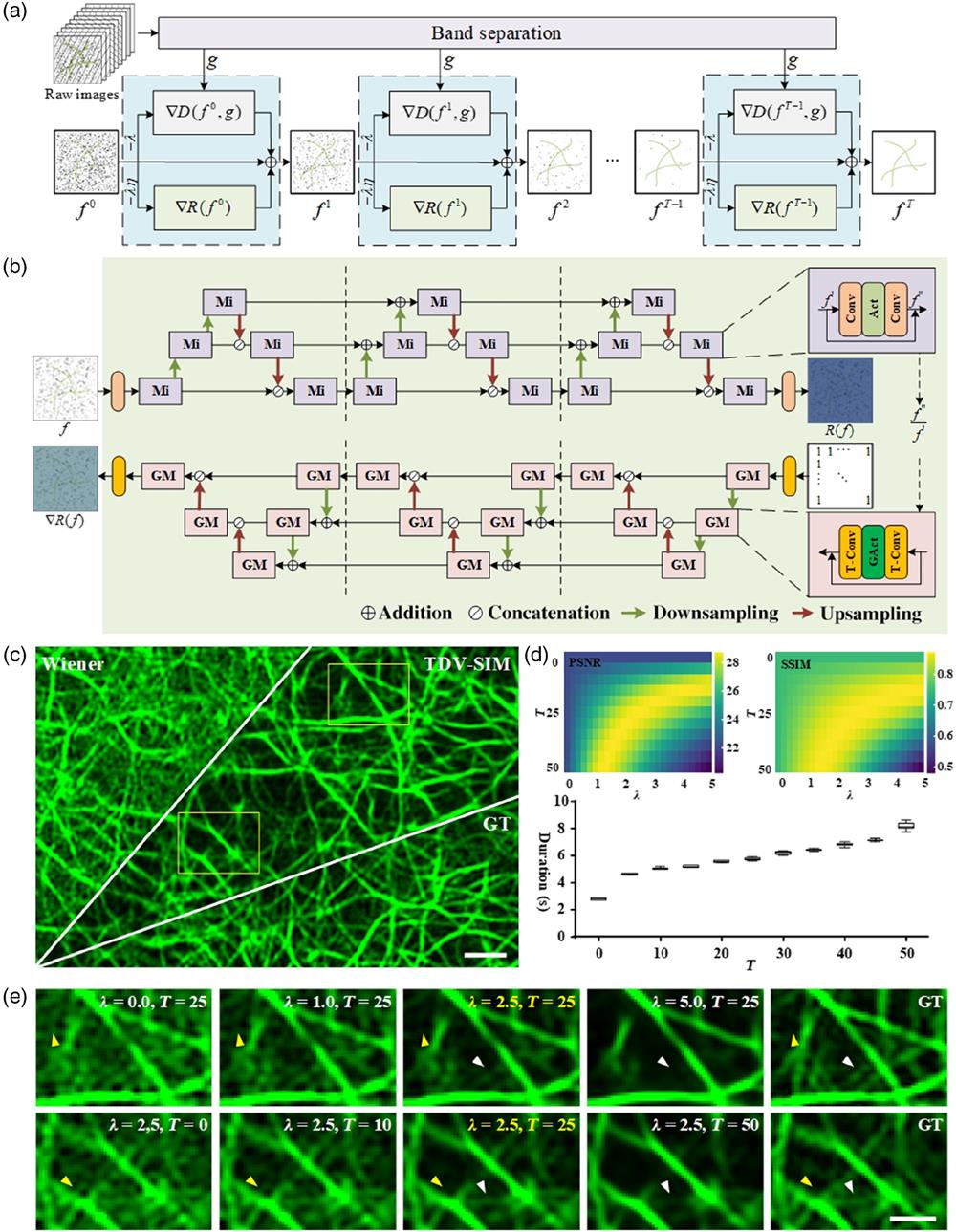
Figure 1.TDV-SIM diagrams and parameter selection. (a) TDV-SIM reconstruction pipeline. (b) Visualization of TDV and its gradient. Mi is a residual structure micro-block and GM is its gradient. Conv is the convolution layer and T-Conv is its gradient. Act is the activation layer and GAct is its gradient. (c) Actin filaments SIM SR image. (d) Top, PSNR and SSIM of TDV-SIM reconstructions with different
Compared to the ground truth (GT) image of actin filaments [averages of multiple Wiener-processed images, Fig. 1(c)], we quantized the peak SNR [PSNR; Fig. 1(d), top left] and structural similarity index measure [SSIM; Fig. 1(d), top right] values of TDV-SIM reconstructions with different weight parameters
2.2 Cell Culture and Labeling
COS-7 cells (ATCC and CRL-1651) were cultured in high-glucose Dulbecco’s modified Eagle’s medium (Gibco, 21063029) supplemented with 10% fetal bovine serum (Gibco) and 1% 100 mM sodium pyruvate solution (Sigma-Aldrich, S8636) in an incubator at 37 °C with 5%
To label mitochondria, COS-7 cells were incubated with 250 nM MitoTracker Green FM (Thermo Fisher Scientific, M7514) in an hank’s balanced salt solution medium (Thermo Fisher Scientific, 14025076) containing
2.3 Image Acquisition, Preprocessing, and Training
The same SIM settings in Hessian-SIM18 were used. To obtain low SNR raw images and the corresponding GT images for training the neural network, we imaged the specimen with SIM. We recorded 20 images for each illumination pattern and then changed the phase and orientation of the pattern. We repeated the cycle nine times, corresponding to three orientations multiplied by three phases, thus obtaining 180 raw images. Then we divided the raw images into 20 groups, with each group containing nine illumination patterns of three phases and three orientations. After removing the fluorescent background, we can obtain 20 SR images with artifacts using Wiener deconvolution. Finally, we mimicked the artifact-free GT by averaging the 20 SR images.
We imaged
2.4 Calculation of Assessment Metrics
To avoid the influence of different methods on the dynamic range of the inferred SR images, we first normalize the SR images,
Artifacts often emerged in regions of minor signals, such as the meshed region within actin filaments. Therefore, benchmarked against the GT, we selected these regions to calculate their variances.
3 Results
3.1 TDV-SIM Excels in Restoring Regular Structures Imaged with a Low SNR
We compared TDV-SIM with other reconstruction methods, including physical-model-based (Wiener deconvolution,11 HiFi-SIM, and Hessian-SIM) and pure DL-based methods [skip-layer connecting U-Nets (scU-Net)24 and deep Fourier channel attention network (DFCAN)25] using synthetic images with known GT (Fig. S1 in the Supplementary Material). TDV-SIM confers balanced performance in generating SR images of high SSIM, low NRMSE, and low artifacts among all reconstruction methods. Next, we examined dynamic actin filaments and ER in live cells observed with short exposures [actin: 1 ms, Fig. 2(a); 2.7 ms, Fig. S2a in the Supplementary Material; and ER: 0.789 ms, Fig. 2(d)]. Despite the improved reconstructions compared to the Wiener deconvolution, HiFi-SIM and Hessian-SIM still produced artifacts due to noise amplification in background regions with low SNR. TDV-SIM produced more continuous actin filaments (Fig. S2e in the Supplementary Material) with fewer artifacts but comparable SSIM values and resolutions to the conventional reconstruction methods [Figs. 2(b), 2(e), and 2(f)–2(j) and Fig. S2d in the Supplementary Material]. In contrast, pure DL-based methods led to reconstruction with fewer artifacts at the price of reduced resolution and decreased SSIM values. In addition, we often observed inaccurate inferences at the intersections of actin filaments and ER [yellow arrows in Figs. 2(c) and 2(e) and Fig. S2c in the Supplementary Material]. Together with the incorrectly inferred actin filaments at regions with extremely low fluorescence intensity (Fig. S3 in the Supplementary Material), these resembled the “hallucination effects” of pure DL methods,27 which was abolished by the TDV-SIM method. Furthermore, we compared TDV-SIM with rationalized DL (rDL) SIM29 on microtubule image from the BioSR25 data set (Fig. S4 in the Supplementary Material). By incorporating prior knowledge of illumination patterns into the DL network, rDL SIM aimed to denoise raw images rationally. Still, it produced punctated artifacts in background regions, which may be suppressed with a notch filter (NF) (white boxed region in Figs. S4(a) and S4(c) in the Supplementary Material). Moreover, we often observed microtubules within densely labeled regions absent from notch-filtered rDL SIM reconstructions (NF-rDL SIM; yellow arrows in Fig. S4b in the Supplementary Material), which was confirmed by the missing spikes in corresponding fluorescence profiles in the bottom. In comparison, TDV-SIM can avoid the missing signal problem of NF-rDL SIM and produce higher-fidelity reconstructions with fewer artifacts and higher SSIM [Figs. S4(b)–S4(d) in the Supplementary Material].
![]()
Figure 2.TDV-SIM outperforms other reconstruction algorithms in suppressing artifacts and hallucinations while maintaining resolution. (a) Actin filaments under the SR-SIM. (b) Magnified views of the larger boxed region in panel (a) reconstructed by Wiener deconvolution, HiFi-SIM, Hessian-SIM, and TDV-SIM. The GT image is shown as the reference. Profiles along the yellow line are on the bottom. (c) Magnified views of the smaller boxed regions in panel (a) reconstructed by Wiener deconvolution, scU-Net, DFCAN, and TDV-SIM. The GT images are shown as references. (d) Time series imaging of ER under the SR-SIM (
3.2 TDV-SIM Enables Better Reconstruction of Intricate Structures Prone to Photobleaching
Photobleaching constitutes a major problem of fluorescence SR imaging, continuously reducing image SNR, and compromising the quality of reconstructed images, especially upon resolving nonstereotypical structures such as mitochondrial cristae.30 Therefore, we benchmarked the performance of TDV-SIM in resolving mitochondrial cristae dynamics for a prolonged time in live cells [Fig. 3(a)]. During the 20 s recording, the fluorescence intensity of MitoTracker decreased by
![]()
Figure 3.TDV-SIM enables accurate reconstruction of intricate and dynamic mitochondrial cristae structures in live cells after prolonged bleaching. (a) Mitochondria under the SR-SIM. (b) Time-dependent bleaching in fluorescence intensities of mitochondria. (c) Magnified views of the larger boxed region in panel (a) reconstructed by scU-Net, DFCAN, and TDV-SIM and the corresponding GT image at 0 s. Profiles along the blue line are on the right. (d) Magnified views of the smaller boxed region in panel (a) reconstructed by Wiener deconvolution, HiFi-SIM, Hessian-SIM, and TDV-SIM and the corresponding GT images at 0, 15, and 20 s. (e) The SSIMs of regions enclosed mitochondria from different reconstructions compared to GT images at 0, 15, and 20 s (
3.3 TDV-SIM Enables Better Reconstruction of Actin Filaments under Nonlinear SIM
In comparison to conventional linear SIM, nonlinear (NL) SIM achieves higher lateral resolution up to
![]()
Figure 4.TDV-SIM enables better reconstruction of actin filaments under NL-SIM. (a) Actin filaments under the NL-SIM. (b), (c) Magnified views of the white boxed regions in panel (a) reconstructed by Wiener deconvolution, Hessian-SIM, DFCAN, and TDV-NL-SIM. The GT image is shown as the reference. Profiles along the yellow line are on the bottom. (d) Magnified views of the yellow boxed regions in panel (a) reconstructed by Wiener deconvolution, Hessian-SIM, DFCAN, and TDV-NL-SIM. The GT image is shown as the reference. Yellow arrowheads indicate the inaccurate reconstructions of pure DL-based methods. (e) Artifact variances of actin filaments from background regions in different reconstructions (
4 Discussion
For traditional reconstruction methods such as Hessian-SIM, the denoising effect is limited to images with a low SNR. In contrast, the pure DL method directly fits the SR image through raw images, in which the fitting process is a black box. Therefore, reconstruction fidelity entirely depends on the network fitting ability and its comparativeness with the sample. For the proposed TDV-SIM, the SR information is extracted from raw images by the conventional frequency-extracting process and then integrated into the TDV network for artifact suppression. By combining the advantages of conventional physical model-based algorithms with DL-based algorithms, TDV-SIM outperforms existing reconstruction methods in removing artifacts associated with regions of low SNR while retaining sharp-contrast intricate structures. For example, the reconstructed actin filaments and ER of TDV-SIM have an 80.1% decrease in the background artifacts compared with Hessian-SIM and a 24.3% increase in signal fidelity compared with DFCAN. Indeed, all current DL-based reconstruction methods generate blurred mitochondrial cristae structures,24,25 highlighting the difficulty of pure data-driven methods in predicting irregular and complicated structures in constant changes. Under such circumstances, incorporating physical constraints about the image formation process becomes critical, as we show here. Therefore, TDV-SIM has significant advantages over pure DL methods in the face of samples with intricate and dynamic structures.
However, the current TDV-SIM has limitations. On the one hand, inherited from conventional restoration algorithms, better reconstruction results depend on choosing ideal parameters. Through comparative experiments, we set the optimal hyperparameters to be 2.5 and 25 for
Starting from a hybrid angle, TDV-SIM presents a novel solution for high-resolution and HiFi SR-SIM reconstruction from low SNR images. Endorsed with reduced photon dosage and associated phototoxicity, improved imaging speed, and extended imaging duration, TDV-SIM will be crucial for SR imaging subcellular structure dynamics in live cells.
Jianyong Wang received his bachelor’s degree in mechanical design, manufacturing, and automation from the University of Electronic Science and Technology of China in 2019 and his master’s degree in software engineering from the School of Software and Microelectronics of Peking University in 2022. His research interest is super-resolution structured illumination microscopy.
Junchao Fan received his bachelor’s and PhD degrees in engineering from Huazhong University of Science and Technology in 2014 and 2020, respectively. He is an associate professor at Chongqing University of Posts and Telecommunications. His research is focused on the imaging processing and reconstruction algorithm of computational imaging.
Bo Zhou received his bachelor’s degree in mechanical design, manufacturing, and automation from Central South University in 2017 and his master’s degree in software engineering from the School of Software and Microelectronics, Peking University in 2020. Currently, he is a PhD student at Cell Secretion and Metabolism Laboratory, Institute of Molecular Medicine, Peking University. His research interests are the reconstruction algorithms of super-resolution fluorescence microscopy.
Xiaoshuai Huang received his bachelor’s degree in science from Wuhan University in 2013 and his PhD from Peking University in 2018. He is an assistant professor at Peking University. From 2018 to 2020, he was trained as a postdoctoral research fellow at Peking University. His research is focused on super-resolution microscopy and cell biology.
Liangyi Chen is a Boya Professor at Peking University. He majored in biomedical engineering as an undergraduate at Xi’an JiaoTong University and a PhD student at Huazhong University of Science and Technology. His lab focused on developing state-of-the-art imaging techniques, including ultrasensitive Hessian structured illumination microscopy, super-resolution fluorescence-assisted diffraction computational tomography, sparse deconvolution enabled mathematical superresolution, and fast high-resolution miniature two-photon microscopy for brain imaging in freely behaving mice. He is also a guest professor at Université PSL and École Normale Supérieure.


Set citation alerts for the article
Please enter your email address

© Copyright 2018-2021 | Chinese Laser Press. All Rights Reserved 沪ICP备15018463号-20