
Cheng Jin, Chi Liu, Lingjie Kong, "High-axial-resolution optical stimulation of neurons in vivo via two-photon optogenetics with speckle-free beaded-ring patterns," Photonics Res. 10, 1367 (2022)
- Photonics Research
- Vol. 10, Issue 6, 1367 (2022)
Abstract
1. INTRODUCTION
In recent decades, optogenetics has played an increasingly important role in probing functional codes of neurons [1–3]. However, in conventional one-photon stimulation based optogenetics, all neurons in the illumination volume are excited or inhibited, which makes it complex to decipher the neural network. To regulate opsins at single-neuron resolutions specifically, two-photon optogenetics has been demonstrated [4,5].
Considering that opsins are distributed on neural membranes, to perform two-photon optogenetics on specific neural ensembles, there are generally two kinds of stimulation strategies: the spiral scan approach [6,7] and scanless holographic approach [8,9]. The former manipulates neurons by spirally scanning somas with diffraction limited foci. This method requires lower laser energy, but may introduce latency and jitter. The latter approach generates extended patterns to cover whole somas based on computer-generated holography (CGH). This method can stimulate neurons without temporal delay caused by scanning, resulting in a better temporal resolution. However, for commonly used extended patterns, such as disks, the axial resolution deteriorates linearly with the increase in radii [10]. Thus, it is difficult to maintain single-neuron resolution, especially when multiple extended patterns are required to simulate neural ensembles simultaneously [11].
To improve the axial resolution of extended patterns in the scanless holographic approach, temporal focusing (TF) technology is adopted [12]. One TF method uses a spatial light modulator (SLM) to generate an extended pattern through the CGH method, and then uses another SLM to achieve three-dimensional (3D) locations of the pattern [13]. Another TF method, called three-dimensional scanless holographic optogenetics with temporal focusing (3D-SHOT), uses a lens [14] or a diffuser [15] to generate an extended pattern followed by using an SLM to position the pattern in 3D. In both methods, because a grating is introduced in a conjugate plane of the focal plane, two-photon absorption is efficient only at the focal plane where the short pulse width recovers. Thus the axial resolution can be effectively improved. However, the introduction of a grating increases the system complexity and the loss of laser power.
Sign up for Photonics Research TOC. Get the latest issue of Photonics Research delivered right to you!Sign up now
In the scanless holographic approach, speckle usually exists when using CGH for extended pattern generation. As the phase of a light field at the target plane is unconstrained in CGH, there are random energy fluctuations on the generated extended patterns, which are more obvious in two-photon optogenetics [16]. To eliminate speckles, various solutions have been proposed. From the perspective of novel CGH algorithms, holographic speckles can be depressed by introducing an initial phase at target planes [17], using a bandwidth constraint strategy [18,19], adopting zero padding [20], and employing deep learning [21]. But in these methods, parameters should be optimized for specific goals, which increases computational complexity. From the perspective of hardware development, time averaging of multiple irrelevant holograms can also improve the uniformity of holograms [22,23], but this method is not applicable in two-photon optogenetics as it sacrifices temporal resolution.
Here, we propose a novel scheme to achieve high-axial-resolution optical stimulation of neurons
2. MATERIALS AND METHODS
A. Optical Setup
We experimentally characterize two-dimensional (2D) intensity distribution of the generated holographic patterns by the vertical detection method [24] with two-photon excited fluorescence in a dye pool of sulforhodamine 101 (CAS#60311-02-6, Shanghai Yuanye Bio-Technology). To test 3D intensity distribution when multiple patterns are generated simultaneously, we build a detection system with opposite-facing objectives (Fig. 1). The device used in the excitation path is equivalent to that of the vertical detection system [24]. The sample is a custom-made thin film of fluorescent paint (Tamiya Color TS-36 Fluorescent Red) on cover glasses. The emission fluorescence is imaged on a CCD (33UX178, DMK) by objective 2 (
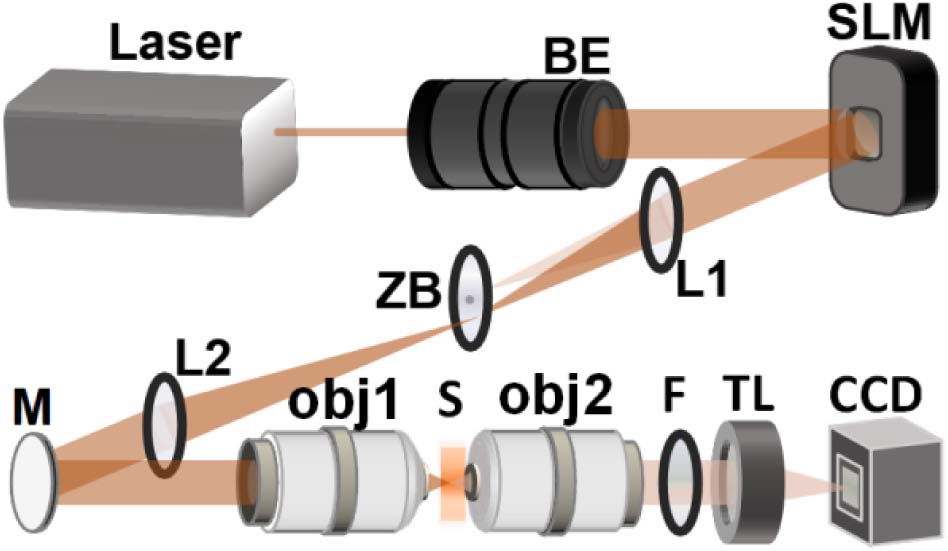
Figure 1.Detection system with opposite-facing objectives. BE, beam expander; ZB, zero-order blocker; L, lens; M, mirror; obj, objective; S, sample; F, filter; TL, tube lens.
To verify the improved performance of our method via two-photon based all-optical physiology
B. System Registration
A hydrogel layer with sulforhodamine 101 is used for the registration between holographic coordinates and imaging coordinates. The weighted Gerchberg-Saxton (GSw) algorithm [25] is used to generate foci of different coordinates at the target imaging plane, and the corresponding phase distribution is loaded onto the SLM. The hydrogel layer is ablated using two-photon stimulation mode with a target phase, and the spatial position of every ablation focus is obtained in two-photon imaging mode. The affine transformation relationship between holographic coordinates and imaging coordinates is calculated with the spatial information of at least three ablation foci. Thus, positions of neurons to be stimulated can be calculated in the SLM coordinate system, and holographic patterns can be generated correspondingly. To improve stimulation accuracy, affine transformation relationships are measured at different depths correspondingly.
C. Holographic Pattern Generation
Holograms are obtained after 100-times iteration with the GSw algorithm [25]. A disk pattern commonly used for optical simulation covers the whole circle region. In this work, two novel patterns are generated and tested, and compared with the disk pattern. One is an annular pattern, and the other is a beaded-ring pattern, which consists of multiple foci evenly spaced along a circle. For annular patterns, the pattern width is 1 μm, which is defined as outer radius minus inner radius. For beaded-ring patterns, eight foci along a circle with radii of 5 μm are used for most experiments, except the patterns in Figs. 2(b)–2(f). In calculating the holographic phase of beaded-ring patterns, the target size of each bead is one pixel, which corresponds to a transform-limited focus, so that the smallest bead size can be achieved.
![]()
Figure 2.Characterization of generated holographic patterns for a single target stimulation. (a)–(f) Two-photon intensity distributions in
![]()
Figure 3.Characterization of generated holographic patterns for multiple target stimulation in a plane. (a)–(f) Two-photon intensity distributions in
In all-optical physiology
D. Sample Preparation and Anesthesia Operation
All procedures involving mice are approved by the Animal Care and Use Committees of Tsinghua University. We label the neurons in mouse L2/3 of S1 cortex via virus infection with a single integrated ChRmine/GCaMP6m virus [26]. Around 300 nL virus is injected at 250 μm depth from the surface of S1 cortex, in 7–8 min. After two weeks of virus expression, chronic optical windows are installed after craniotomy [27]. Then after one week’s recovery, all-optical physiology tests are performed with the mice being anesthetized with isoflurane, using a small animal gas anesthesia machine (R520IP, RWD Life Science).
E. Data Processing
In fluorescence intensity characterization experiments, we integrate intensity within the target pattern at each
To process data in all-optical physiology experiments, a customized MATLAB code is used. After summing two-photon signals in a target neuron region and removing background near the neuron, the calcium signal response
3. RESULTS
A. 2D Performance Characterization of Holographic Patterns
We first use a vertical detection system for the characterization of a single holographic pattern. For a beaded-ring pattern, when eight foci equally spaced along the circle with radii of 5 μm are generated, the average lateral resolution of these foci is 1.5 μm [Figs. 2(a) and 2(g)], while the axial resolution is
Next, we characterize multiple holographic patterns with the vertical detection system. The intensity distributions of multiple patterns generated on the same lateral and axial plane by different schemes are shown in Figs. 3(a)–3(c) and Figs. 3(d)–3(f). As shown in Figs. 3(g) and 3(h), higher axial resolution can be achieved with beaded-ring patterns, for both lateral and axial multiple pattern generation. Especially, when multiple targets are generated on different axial planes, part of the energy is distributed in non-targeted areas for disk patterns [Fig. 3(f)] and annular patterns [Fig. 3(e)]. Significantly, this phenomenon can be avoided in the case of beaded-ring patterns [Fig. 3(d)].
B. 3D Performance Characterization of Holographic Patterns
We further characterize multiple holographic patterns in 3D [Figs. 4(a)–4(c)], using a detection system with opposite-facing objectives, shown in Fig. 1. The GSw algorithm is used to generate 10 targets of beaded-ring, annular, and disk patterns. They all have radii of 5 μm and are distributed in a 3D space of
![]()
Figure 4.Characterization of generated holographic patterns for multiple target stimulation in 3D stack. (a)–(c) Intensity distributions (upper) and maximum intensity projections in
C. All-Optical Physiology
To verify the performance enhancement of our proposed beaded-ring patterns for two-photon optogenetics
![]()
Figure 5.All-optical physiology test with different stimulation patterns on a single neuron
We achieve the location of neurons with two-photon imaging mode. For a target neuron in an imaging field of view [Fig. 5(b)], two-photon optogenetics based on disk and beaded-ring patterns is employed to stimulate the neuron, and duration is 100 ms for each stimulation. Here, a long duration is chosen to improve responses of neurons and avoid the signal drowning in noise, as it decreases when stimulation patterns move away from neurons along
The stimulation depth where the maximum
Next, we perform two-photon optogenetics stimulation and two-photon calcium signal imaging experiments on multiple neurons simultaneously. As shown in Fig. 6(a) left, two neurons to be stimulated are selected in the same field of view, limited by the available laser power. Then, beaded-ring and disk patterns matching these two neurons are used for stimulation [Fig. 6(a) right], with duration of 100 ms. Two neurons stimulated at the same time are defined as a group. Compared with disk patterns, the average axial resolution of neurons in a group stimulated by beaded-ring patterns is increased by 24.27% [Fig. 6(b)], from
![]()
Figure 6.All-optical physiology test with different stimulation patterns on multiple neurons simultaneously
4. DISCUSSION AND CONCLUSION
Considering that opsins are distributed on the cellular membrane, using a beaded-ring pattern focused on the membrane allows the photostimulation of opsins more efficiently. However, the responses of neurons depend not only on the photostimulation intensity on opsins, but also on the overall illumination volumes of opsins. We test the responses of neurons with stimulations of beaded-ring and disk patterns at different powers (Fig. 7). With the increase in stimulation power, neural responses tend to be saturated under both stimulation patterns. There is no significant difference in neural responses induced by beaded-ring and disk patterns at the same stimulation power.
![]()
Figure 7.Calcium signal of neurons with different types of stimulation patterns, at different stimulation powers. BR, beaded-ring pattern; D, disk pattern; ns, not significant.
In this paper, we propose a novel speckle-free, beaded-ring pattern for holographic two-photon optogenetics
Acknowledgment
Acknowledgment. The authors thank Yang Lin and Kuikui Fan for help with bio-sample preparation. LK acknowledges the support from Tsinghua University.
References
[1] E. S. Boyden, F. Zhang, E. Bamberg, G. Nagel, K. Deisseroth. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci., 8, 1263-1268(2005).
[2] V. Szabo, C. Ventalon, V. De Sars, J. Bradley, V. Emiliani. Spatially selective holographic photoactivation and functional fluorescence imaging in freely behaving mice with a fiberscope. Neuron, 84, 1157-1169(2014).
[3] K. Daie, K. Svoboda, S. Druckmann. Targeted photostimulation uncovers circuit motifs supporting short-term memory. Nat. Neurosci., 24, 259-265(2021).
[4] V. Nikolenko, K. E. Poskanzer, R. Yuste. Two-photon photostimulation and imaging of neural circuits. Nat. Methods, 4, 943-950(2007).
[5] H. Adesnik, L. Abdeladim. Probing neural codes with two-photon holographic optogenetics. Nat. Neurosci., 24, 1356-1366(2021).
[6] W. Yang, L. Carrillo-Reid, Y. Bando, D. S. Peterka, R. Yuste. Simultaneous two-photon imaging and two-photon optogenetics of cortical circuits in three dimensions. eLife, 7, e32671(2018).
[7] R. Prakash, O. Yizhar, B. Grewe, C. Ramakrishnan, N. Wang, I. Goshen, A. M. Packer, D. S. Peterka, R. Yuste, M. J. Schnitzer, K. Deisseroth. Two-photon optogenetic toolbox for fast inhibition, excitation and bistable modulation. Nat. Methods, 9, 1171-1179(2012).
[8] A. Forli, D. Vecchia, N. Binini, F. Succol, S. Bovetti, C. Moretti, F. Nespoli, M. Mahn, C. A. Baker, M. M. Bolton, O. Yizhar. Two-photon bidirectional control and imaging of neuronal excitability with high spatial resolution
[9] J. V. Gill, G. M. Lerman, H. Zhao, B. J. Stetler, D. Rinberg, S. Shoham. Precise holographic manipulation of olfactory circuits reveals coding features determining perceptual detection. Neuron, 108, 382-393(2020).
[10] C. Lutz, T. S. Otis, V. DeSars, S. Charpak, D. A. DiGregorio, V. Emiliani. Holographic photolysis of caged neurotransmitters. Nat. Methods, 5, 821-827(2008).
[11] E. Papagiakoumou, E. Ronzitti, I. W. Chen, M. Gajowa, A. Picot, V. Emiliani. Two-photon optogenetics by computer-generated holography. Optogenetics: A Roadmap, 175-197(2018).
[12] E. Papagiakoumou, V. De Sars, D. Oron, V. Emiliani. Patterned two-photon illumination by spatiotemporal shaping of ultrashort pulses. Opt. Express, 16, 22039-22047(2008).
[13] O. Hernandez, E. Papagiakoumou, D. Tanese, K. Fidelin, C. Wyart, V. Emiliani. Three-dimensional spatiotemporal focusing of holographic patterns. Nat. Commun., 7, 11928(2016).
[14] N. C. Pégard, A. R. Mardinly, I. A. Oldenburg, S. Sridharan, L. Waller, H. Adesnik. Three-dimensional scanless holographic optogenetics with temporal focusing (3D-SHOT). Nat. Commun., 8, 1(2017).
[15] G. J. Goldey, D. K. Roumis, L. L. Glickfeld, A. M. Kerlin, R. C. Reid, V. Bonin, D. P. Schafer, M. L. Andermann. Removable cranial windows for long-term imaging in awake mice. Nat. Protoc., 9, 2515-2538(2014).
[16] E. Ronzitti, C. Ventalon, M. Canepari, B. C. Forget, E. Papagiakoumou, V. Emiliani. Recent advances in patterned photostimulation for optogenetics. J. Opt., 19, 113001(2017).
[17] H. Pang, J. Wang, A. Cao, Q. Deng. High-accuracy method for holographic image projection with suppressed speckle noise. Opt. Express, 24, 22766-22776(2016).
[18] L. Chen, S. Tian, H. Zhang, L. Cao, G. Jin. Phase hologram optimization with bandwidth constraint strategy for speckle-free optical reconstruction. Opt. Express, 29, 11645-11663(2021).
[19] X. Sui, Z. He, G. Jin, D. Chu, L. Cao. Band-limited double-phase method for enhancing image sharpness in complex modulated computer-generated holograms. Opt. Express, 29, 2597-2612(2021).
[20] W. Qu, H. Gu, Q. Tan. Holographic projection with higher image quality. Opt. Express, 24, 19179-19184(2016).
[21] D. Y. Park, J. H. Park. Hologram conversion for speckle free reconstruction using light field extraction and deep learning. Opt. Express, 28, 5393-5409(2020).
[22] J. Amako, H. Miura, T. Sonehara. Speckle-noise reduction on kinoform reconstruction using a phase-only spatial light modulator. Appl. Opt., 34, 3165-3171(1995).
[23] L. Golan, S. Shoham. Speckle elimination using shift averaging in high-rate holographic projection. Opt. Express, 17, 1330-1339(2009).
[24] C. Jin, C. Liu, R. Shi, L. Kong. Precise 3D computer-generated holography based on non-convex optimization with spherical aberration compensation (SAC-NOVO) for two-photon optogenetics. Opt. Express, 29, 20795-20807(2021).
[25] R. Di Leonardo, F. Ianni, G. Ruocco. Computer generation of optimal holograms for optical trap arrays. Opt. Express, 15, 1913-1922(2007).
[26] J. H. Marshel, Y. S. Kim, T. A. Machado, S. Quirin, B. Benson, J. Kadmon, C. Raja, A. Chibukhchyan, C. Ramakrishnan, M. Inoue, J. C. Shane. Cortical layer–specific critical dynamics triggering perception. Science, 365, 5202(2019).
[27] A. R. Mardinly, I. A. Oldenburg, N. C. Pégard, S. Sridharan, E. H. Lyall, K. Chesnov, S. G. Brohawn, L. Waller, H. Adesnik. Precise multimodal optical control of neural ensemble activity. Nat. Neurosci., 21, 881-893(2018).


Set citation alerts for the article
Please enter your email address

© Copyright 2018-2021 | Chinese Laser Press. All Rights Reserved 沪ICP备15018463号-20