
W Pearson MATTHEW. A Case Study of Allocasuarina robusta Recovery Using History and Biogeography to Identify Future Priorities[J]. Journal of Resources and Ecology, 2020, 11(2): 171
- Journal of Resources and Ecology
- Vol. 11, Issue 2, 171 (2020)
Abstract
1 Introduction
Lindenmayer (2019) synopsis, challenges our concepts towards the size of conservation areas and what size means towards conservation. The synopsis adds to the debate of single large or serval small areas for conservation (
Pilgrim et al. (2004) suggest viable populations of trees, as opposed to shrubs, require an extensive range due to pollination requirements. The type of pollination which Pilgrim et al. (2004) identified for trees was predominantly wind. Understanding population dynamics is seen as crucial to allow the conservation category to be validated (
Species classification within a conservation category follows a continuum from critically endangered to least concern. Deciding where to place a species along this continuum is sometimes based on a species spatial distribution (
2 The Study species - Allocasuarina robusta
Pollination in members of Casuarinaceae occurs by the wind; the bracteoles develop into a fruit that contains a single winged samara seed (
The conservation status of
3 Allocasuarina robusta Botanical data
An
One of the processes used for defining flora for conservation is using specimens collected and then using the specimens to generate a model of what makes a species rare (
![]()
Figure 1.Map of
Understanding the environmental factors impacting on flora may aid conservation practices, but it might not conserve a specific species (
Given and Norton (1993) concluded that a species must be one which has widespread appeal with the public. How this process identifies which species to protect or reinforce Given and Norton (1993) questioned. The natural extension of Given and Norton (1993) is the development of an action plan describing the priorities for conserving a species. These schemes work well within government, although they are considered flawed through thinking of each species on a linear continuum (
Population modelling needs to use a suitably sized population to enable a species to be self-sustaining. Quarmby (2011) validated populations of
4 Supportive or complementary data sources
Without this data, the next best option is to use historical land use accounts (
By the 1870’s cropping in the flat plateau areas, had concluded which incorporates the Hindmarsh Tiers area (
By the 1920s, extensive clearing for agriculture occurred around Victor Harbor (
Herbarium data does not provide insight into the type of land management practices for facilitating a species recovery. In the case of
The loss of vegetation via European colonisation on the Fleurieu Peninsula is estimated to be as high as 42% (
Paleontological records of Holocene sediments show
Fire can alter a vegetation community, causing lasting impacts for decades and possibly centuries to come (
A balance needs to occur between managing fire to maintain biodiversity and the risk of unpredictable fires impacting on human activities (
5 Relating to Allocasuarina robusta
Relating these land management practices and observation to
Environmental disturbance represents the most significant impact on Australian flora (
The lesson learnt from a species recovery and regeneration from changed natural processes could be applied to rare or threatened species, i.e.
The physical floristic specimen can provide genetic information, another source of evidence is the information on the specimen card provided by the collector (
![]()
Figure 2.Map of
Removal of
The supposition from biogeography influences on distribution may be an extension of the investigation conducted by
Biogeographical investigations from
The population data on
![]()
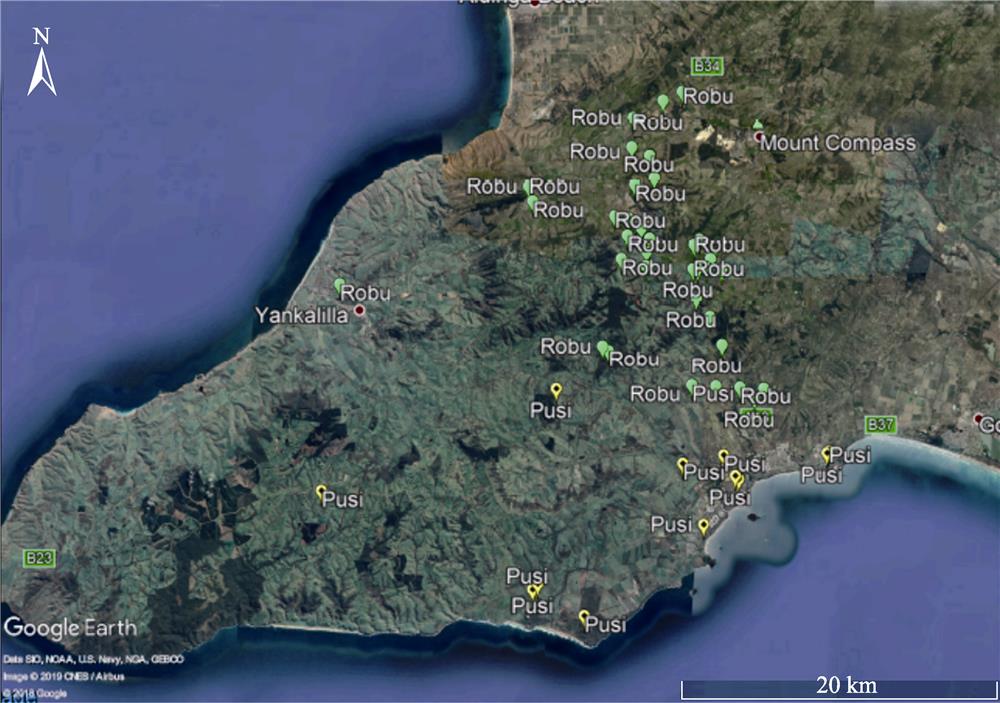
Figure 3.Zoomed in to a section of
Note: Robu =
![]()
Figure 4.A zoomed-in section of
Note: Map created in Open Source ESRI GIS.
![]()
Figure 5.Edited version of
Geographical gradients can impact on species morphology, while a revision on
To increase genetic diversity, Quarmby (2011) took remedial action by reinforcing populations with seedlings from other metapopulations. Quarmby (2011) remedial actions were using the best intentions but not knowing at a genetic level how each fragment of
To highlight the importance of understanding a species breeding system would be as Collier and Garnett (2018) describes the difference which can occur in sowing seeds by collecting seeds leading to the demise of the parent plant. Collier and Garnett (2018) acknowledge sowing seeds and the relationship seeds have with fire is not well understood. The size of populations can increase the risks posed to extinction either at various scales locally, regional or statewide (
6 Moving forward
As a society, the process of classifying threatened species can have significant implications. In the case of
The breeding systems in the context of fragmentation can provide a vital process into defining the needs to protect a species/population or better manage an existing population of
References
[1] Bardsley DK, Sweeney SM. Guiding climate change adaptation within vulnerable natural resource management systems. Environmental Management, 45, 1127-1141(2010).
[2] Bardsley DK, WeberD, Robinson GM et al. Wildfire risk, biodiversity and peri-urban planning in the Mt Lofty Ranges, South Australia. Applied Geography, 63, 155-165(2015).
[3] BickfordS, GellP. Holocene vegetation change, Aboriginal wetland use and the impact of European settlement on the Fleurieu Peninsula, South Australia. The Holocene, 15, 200-215(2005).
[4] BickfordS, GellP, Hancock GJ. Wetland and terrestrial vegetation change since European settlement on the Fleurieu Peninsula, South Australia. The Holocene, 18, 425-436(2008).
[5] BickfordS, MackeyB. Reconstructing pre-impact vegetation cover in modified landscapes using environmental modelling, historical surveys and remnant vegetation data: A case study in the Fleurieu Peninsula, South Australia. Journal of Biogeography, 31, 787-805(2004).
[6] Bradstock RA, Keith DA, Auld TD. 1995. Fire conservation: Imperatives and constraints on managing for diversity. In: Bradstock R A, Auld T D, Keith D A, et al. Conservation biodiversity: Threats and solutions. Chipping Borton: Surrey Beatty and Sons:, 323-333.
[7] Callister KE, Florentine SK, Westbrooke ME. An investigation of the soil seedbank and seed germination of perennial species in Belah (. Casuarina pauper) woodlands in north-west Victoria. Australian Journal of Botany, 66, 202-212(2018).
[8] ClarkeB, RuddD, HarveyN. The ‘sea change’ phenomenon in South Australia. South Australia Geographical Journal, 107, 69-85(2008).
[9] CollierP, GarnettR. Threatened plant translocation case study:. Wrinkled Dollybush), Asteraceae. Australasian Plant Conservation, 26, 21-24(2018).
[10] Corlett RT. Plant diversity in a changing world: Status, trends, and conservation needs. Plant Divers, 38, 10-16(2016).
[11] Crossman ND, Bryan BA, Cooke DA. An invasive plant and climate change threat index for weed risk management: Integrating habitat distribution pattern and dispersal process. Ecological Indicators, 11, 183-198(2011).
[12] Dilcher DL, Christophel DC, Bhagwandin HO et al. Evolution of the. Casuarinaceae: Morphological comparisons of some extant species. American Journal of Botany, 77, 338-355(1990).
[13] Dolan RW, Moore ME, Stephens JD. Documenting effects of urbanization on flora using herbarium records. Journal of Ecology, 99, 1055-1062(2011).
[14] Elzinga CL, Salzer DW, Willoughby JW. Measuring & monitoring plant populations. Denver Colorado: Bureau of. Land Management National Business Center.(1998).
[15] Emery NJ, HillierP, Offord CA. Threatened plant translocation case study:. North Rothbury Persoonia), Proteaceae. Australasian Plant Conservation, 26, 18-21(2018).
[16] Farrel TP, Ashton DH. Population studies on. Acacia melanoxylon R. Br. I. variation in seed and vegetative characteristics. Australian Journal of Botany, 26, 365-379(1978).
[17] FordT. Population change in Adelaide’s Peri-urban region: Patterns, causes and implication. Master diss., University of. Adelaide.(1998).
[18] Fusco DA, McDowell MC, Prideaux GJ. Late-Holocene mammal fauna from southern Australia reveals rapid species declines post-European settlement: Implications for conservation biology. The Holocene, 26, 699-708(2015).
[19] Gaston KJ. Valuing common species. Science, 327, 154-155(2010).
[20] Given DR, Norton DA. A multivariate approach to assessing threat and for priority setting in threatened species conservation. Biological Conservation, 64, 57-66(1993).
[21] Gogol-ProkuratM. Predicting habitat suitability for rare plants at local spatial scales using a species distribution model. Ecological Applications, 21, 33-47(2011).
[22] Guerin GR. Monitoring diversity and tree demography in vegetation communities of the Mount Lofty - Flinders Ranges. The South Australian Naturalist, 91, 56-67(2017).
[24] HansenM.J, Clevenger AP. The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biological Conservation, 125, 249-259(2005).
[25] Harding CL. 4.3 Threatening processes and management issues. Wetland Inventory for the Fleurieu Peninsula. Harding C L. Adelaide South Australia, National Action Plan for Salinity and Water Quality / Department for Environment &. Heritage.(2005).
[26] Heywooda VH, Iriondo JM. Plant conservation: Old problems, new perspectives. Biological Conservation, 113, 321-335(2003).
[27] JessopJ, Black JM, Toelken HR. Flora of South Australia: Lycopodiaceae-Rosaceae. South Australian Government Printing Division.(1986).
[28] Johnson L AS. Notes on. Casuarinaceae II. Journal of Adelaide Botanic Gardens, 6, 73-87(1982).
[29] KeanJ, BarlowN. Exploring rarity using a general model for distribution and abundance. The American Naturalist, 163, 407-416(2004).
[30] Kricsfalusy VV, TrevisanN. Prioritizing regionally rare plant species for conservation using herbarium data. Biodiversity and Conservation, 23, 39-61(2013).
[31] Ladiges PY, Bayly MJ, NelsonG. Searching for ancestral areas and artifactual centers of origin in biogeography: With comment on east-west patterns across southern Australia. Systematic Biology, 61, 703-708(2012).
[32] LindenmayerD. Small patches make critical contributions to biodiversity conservation. Proceedings of the National Academy of Sciences, 116, 717-719(2019).
[33] Lopez Ortiz MJ, MarceyT, Lucash MS et al. Post-fire management affects species composition but not Douglas-fir regeneration in the Klamath Mountains. Forest Ecology and Management, 432, 1030-1040(2019).
[34] LothianA, HarrisC. Clearance of native vegetation in the Mount Lofty Ranges 1945-68. South Australian Geographical Journal, 113, 29-42(2014).
[35] Lunt ID, Spooner PG. Using historical ecology to understand patterns of biodiversity in fragmented agricultural landscapes. Journal of Biogeography, 32, 1859-1873(2005).
[36] Master LL. Assessing threats and setting priorities for conservation. Conservation Biology, 5, 559-563(1991).
[37] McIntyreS, LavorelS. Predicting richness of native, rare, and exotic plants in response to habitat and disturbance variables across a variegated landscape. Conservation Biology, 8, 521-531(1994).
[38] MorganJ. Have tube stock plantings successfully established populations of rare grassland species into reintroduction sites in western Victoria?. Biological Conservation, 89, 235-243(1999).
[39] Morrison DA. Effects of fire intensity on plant species composition of sandstone communities in the Sydney region. Austral Ecology, 27, 433-441(2002).
[40] MotzkinG, WilsonP, Foster DR et al. Vegetation patterns in heterogeneous landscapes: The importance of history and environment. Journal of Vegetation Science, 10, 903-920(1999).
[41] NualartN, Ibá?ezN, SorianoI et al. Assessing the relevance of herbarium collections as tools for conservation biology. The Botanical Review, 83, 303-325(2017).
[42] Ottewell KM, Bickerton DC, ByrneM et al. Bridging the gap: A genetic assessment framework for population-level threatened plant conservation prioritisation and decision-making. Diversity and Distributions, 22, 174-188(2016).
[43] Pannell JR, Myerscough PJ. Canopy-stored seed banks of. Allocasuarina distyla and A. nana in relation to time since fire. Australian Journal of Botany, 41, 1-9(1993).
[44] PatonP, CromptonJ. Birds of the hundred of Encounter Bay listed by John W Crompton from 1909 to 1925. South Australian Ornithologist, 39, 18-35(2013).
[45] Pilgrim ES, Crawley MJ, DolphinK. Patterns of rarity in the native British flora. Biological Conservation, 120, 161-170(2004).
[46] Pressey RL, Nicholls AO. Efficiency in conservation evaluation: Scoring versus iterative approaches. Biological Conservation, 50, 199-218(1989).
[47] QuarmbyJ. Action plan for mount compass oak--Bush (
[48] ReuterD. The South Australian superphosphate story - Part I. Agricultural Science, 1, 23-33(2012).
[49] Rivers MC, TaylorL, Brummitt NA et al. How many herbarium specimens are needed to detect threatened species?. Biological Conservation, 144, 2541-2547(2011).
[50] Roxburgh SH, SheaK, Wilson JB. The intermediate disturbance hypothesis: Patch dynamics and mechanisms of species coexistence. Ecology, 85, 359-371(2004).
[51] SalzerD, SalafskyN. Allocating Resources Between Taking Action, Assessing Status, and Measuring Effectiveness of Conservation Actions. Natural Areas Journal, 26, 310-316(2006).
[52] Spooner PG, Lunt ID. The influence of land-use history on roadside conservation values in an Australian agricultural landscape. Australian Journal of Botany, 52, 445-458(2004).
[53] Steane DA, Wilson KL, Hill RS. Using matK sequence data to unravel the phylogeny of Casuarinaceae. Molecular Phylogenetics and Evolution, 28, 47-59(2003).
[54] Stenhouse RN. Assessing disturbance and vegetation condition in urban bushlands. Australasian Journal of Environmental Management, 12, 16-26(2005).
[55] Swamy B GL. A contribution to the life history of
[56] Swetnam TW, Allen CD, Betancourt JL. Applied historical ecology: Using the past to manage for the future. Ecological Applications, 9, 1189-1206(1999).
[57] TurnbullJ. Historical GIS - Examples from the Creswick Goldfields. Geospatial science research,. CEUR workshop proceedings, 2, 37-46(2012).
[58] ViolleC, ThuillerW, MouquetN et al. Functional rarity: The ecology of outliers. Trends of Ecology and Evolution, 32, 356-367(2017).
[59] Williams JN, SeoC, ThorneJ et al. Using species distribution models to predict new occurrences for rare plants. Diversity and Distributions, 15, 565-576(2009).
[60] Willis CG, Ellwood ER, Primack RB et al. Old plants, new tricks: Phenological research using herbarium specimens. Trends of Ecology and Evolution, 32, 531-546(2017).
[62] WilsonK. Johnson L A S. 1989. Casuarinaceae. In: Robertson R et al., eds. Flora of Australia, George A S. Canberra ACT Australia, Australian Government Printing Service:, 3, 100-174.


Set citation alerts for the article
Please enter your email address

© Copyright 2018-2021 | Chinese Laser Press. All Rights Reserved 沪ICP备15018463号-20