
Wei GAO, Meimei LIN, Yongrong HUANG, Shide HUANG, Gongfu YE, Zhiqun HUANG. Effects of Forest Types and Environmental Factors on Soil Microbial Biomass in a Coastal Sand Dune of Subtropical China[J]. Journal of Resources and Ecology, 2020, 11(5): 454
- Journal of Resources and Ecology
- Vol. 11, Issue 5, 454 (2020)
Abstract
Keywords
1 Introduction
Soil microorganisms play important roles in soil carbon (C) and nitrogen (N) cycling and mineralization, and they are highly sensitive to changes in soil conditions (
The metabolic processes of soil microorganisms are affected by the comprehensive characteristics of plant community, soil temperature, soil moisture, soil pH, soil foundation fertility, and substrate (
Sand dunes are common on the southeastern coast of China, where the infertile soil has low water-holding capacity and high salinity. Casuarina equisetifolia L. is an actinorhizal N-fixing species with good wind and salt resistance, and it is often used for the establishment of protection forests in coastal sandy areas. C. equisetifolia plantations in China cover approximately 3×105 ha and play an important role in local environment improvement and ecological security maintenance efforts (
In coastal sand dunes, the knowledge of environmental controls and variations in their soil microbial biomass across forest types in coastal sandy areas will help to guide afforestation species selection and ecosystem management. Therefore, it is important to understand how the differences in species composition and the seasonal fluctuations of soil temperature and moisture can affect the dynamics of soil microbial biomass, and what roles soil microbial biomass play in soil C and N cycling. The goals of this study were to: 1) Investigate the soil MBC and MBN contents and their seasonal dynamics across five forest types (a secondary forest of L. glutinosa and plantations of Casuarinas, Pine, Acacia, and Eucalyptus) in the southeastern coast of Fujian province; 2) Identify factors that correlate with soil MBC and MBN contents, including litterfall, litter C and N contents, fine root biomass, and root C and N contents; and 3) Measure the physical and chemical properties of the soils, including soil temperature and soil moisture; and evaluate their effects on the soil C and N cycling across the different forest types.
2 Materials and methods
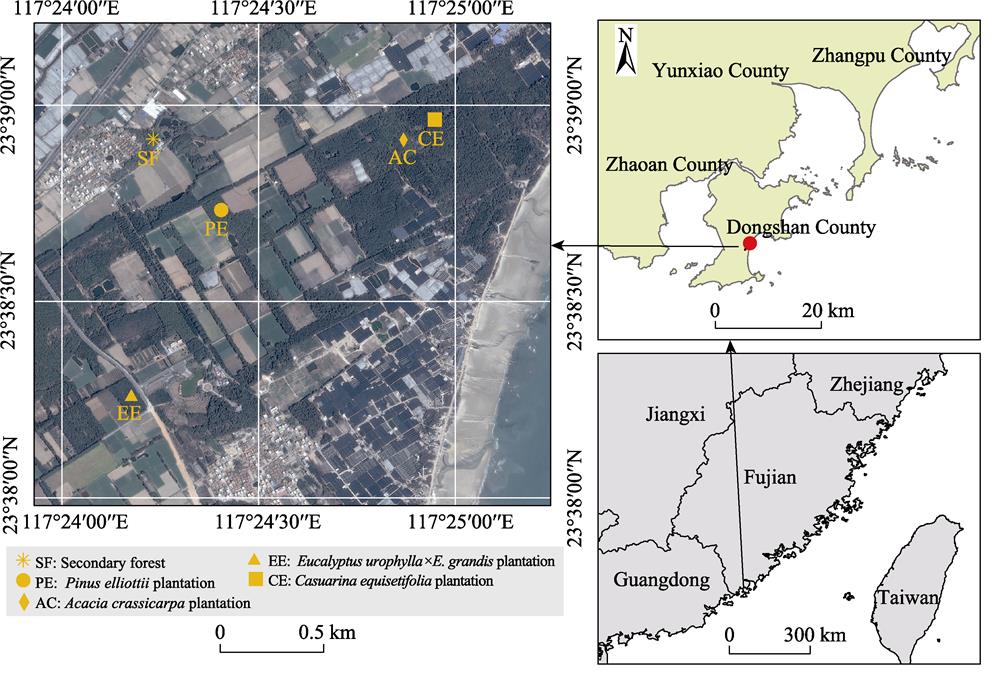
2.1 Study site
The study was conducted at Chishan Forestry Center of Dongshan County (23°38°N, 117°24°E) in southeastern Fujian Province, China (
![]()
Figure 1.Fig. 1
![]()
Figure 2.Fig. 2
Five forest stands, including a secondary forest and plantations of Casuarina, Pine, Acacia, and Eucalyptus, were selected as experimental groups in December 2014. These stands have similar elevations and soil types, each with an average slope of less than 10°. The detailed site characteristics and topsoil properties of the five forest stands are shown in
| Variable | Secondary forest | ||||
|---|---|---|---|---|---|
| Stand age (yr) | 11 | 21 | 22 | 22 | >50 |
| Forest average DBH (cm) | 15.60 | 21.00 | 21.10 | 17.90 | 9.70 |
| Mean tree height (m) | 11.40 | 13.90 | 12.50 | 14.90 | 5.60 |
| Tree density (ind ha-1) | 1300 | 1500 | 950 | 1600 | 1400 |
| Litterfall (mg ha-1 yr-1) | 12.32ab | 12.99a | 10.06b | 10.08b | 12.67a |
| Litter C (g kg-1) | 473.40bc | 499.70ab | 513.60a | 497.70ab | 495.80c |
| Litter N (g kg-1) | 9.21c | 6.58d | 15.10b | 8.83c | 16.41a |
| Litter C:N ratio | 51.55b | 77.12a | 34.07c | 56.43b | 28.05c |
| Litter cellulose (g kg-1) | 114.80d | 217.90b | 172.10c | 275.20a | 157.40c |
| Litter lignin (g kg-1) | 179.90e | 365.90b | 449.30a | 227.50d | 300.60c |
| Litter lignin: N ratio | 19.60c | 56.50a | 29.80b | 25.80b | 18.30c |
| Root Biomass (mg ha-1) | 2.16ab | 1.05b | 1.84ab | 3.04a | 2.77a |
| Root C (g kg-1) | 376.40ab | 365.10b | 304.80c | 412.70a | 378.00ab |
| Root N (g kg-1) | 5.70c | 6.30c | 10.50b | 19.70a | 19.20a |
| Root C: N ratio | 69.90a | 63.50a | 29.00b | 20.90b | 19.80b |
| Bulk density (g cm-3) | 1.36b | 1.23cd | 1.28bc | 1.45a | 1.17d |
| pH | 5.00b | 4.65c | 4.73c | 4.71c | 6.57a |
| Total C (g kg-1) | 6.65b | 4.33b | 4.08b | 3.17b | 12.71a |
| Total N (g kg-1) | 0.54b | 0.53bc | 0.53bc | 0.27c | 1.26a |
| Soil C: N ratio | 11.97a | 8.12b | 7.70b | 11.58a | 9.91ab |
| Soil NH4+-N concentration (μg g-1) | 3.52c | 3.78c | 6.27b | 4.38c | 7.00a |
| Soil NO3--N concentration (μg g-1) | 0.20b | 1.14b | 1.47b | 0.87b | 18.20a |
| Total inorganic N (μg g-1) | 2.30b | 3.90b | 6.50b | 3.10b | 25.80a |
| Soil DOC (mg kg-1) | 56.20b | 40.40b | 42.20b | 53.30b | 84.80a |
| Soil DON (mg kg-1) | 2.80bc | 3.30bc | 6.80b | 0.60c | 22.5a0 |
| Available P (mg kg-1) | 0.57b | 0.61b | 1.26b | 0.89b | 20.49a |
| Exchangeable Mg (mg kg-1) | 0.28b | 0.33b | 0.26b | 0.32b | 0.71a |
Table 1.
Detailed site characteristics and topsoil properties of the five forest stands on the southeastern coast of China
The secondary forest is composed of native vegetation resulting from secondary succession with distinct tree, shrub, and herbaceous layers, which regenerates naturally and has been afforded long term protection by local people. The age of the secondary forest is more than 50 years, and the dominant species in the canopy are L. glutinosa and Celtis sinensis Willd. The shrub layer is predominantly Glochidion eriocarpum, Litsea rotundifolia, Bridelia tomentosa, and Anodendron affine (Hook. et Arn.) Druce, and the main herb species are Ophiopogon bodinieri and Arthraxon hispidus var. hispidus. Both Casuarinas and Acacia plantations were established in 1992 with main tree species of C. equisetifolia and Acacia crassicarpa, respectively. The pine plantation was planted with Pinus elliottii Engelm. and slash pine in 1993. The Eucalyptus plantation was established in 2004 with Eucalyptus urophylla × E.grandis crosses; the previous species on this site was P. elliottii, which was planted in 1976. The canopy densities of the different plantations are greater than 0.7 with few shrubs and grasses in the understory (
2.2 Experimental design and measurement
In each of the five forest stands, four 20 m × 20 m plots were established with a 10 m buffer between each plot. In July 2015, ten soil cores (1.0 m length × 5 cm diameter) were collected along the diagonal of each plot from the soil surface to a 100-cm depth. Fine roots (< 2 mm diameter) were washed out and the remaining fine-root biomass was estimated following
Five litter traps (1 m × 1 m) with 1 mm nylon mesh were randomly arranged 0.3 m above the forest floor in each plot for litter collection. From March 2015 to February 2016, litter was collected monthly for determining annual litterfall.
Ten soil pits along the diagonal of each plot were used to collect soil at both 0-10 cm (topsoil) and 10-20 cm (subsoil) depths in April (spring), July (summer), September (autumn), and November (winter) 2015. A 500 g sample was collected from each layer of every plot, and 300 g portions of each were air-dried, ground, and sieved through a 2-mm sieve for microbial biomass analysis. The remaining soil was ground and sieved through a 0.149-mm sieve for total soil carbon (TC) and total N (TN) analysis. Soil bulk density in each layer was calculated using intact soil cores (100 cm3). Soil temperature was obtained using an instantaneous digital thermometer (AM-11T, Avalon company, USA), and soil moisture was determined using an oven-drying method.
Soil microbial biomass C (MBC) and N (MBN) were analyzed following the chloroform fumigation-extraction method (
2.3 Data analysis
Data from each of the four plots were averaged for further analysis. To test the significances of differences in soil C and N contents, soil pH, soil bulk density, as well as the soil MBC and MBN contents between different forest types, we used one-way analysis of variance (ANOVA) with Duncan’s multiple comparison method at a significance level of 0.05. We also used Pearson correlation analysis to determine how annual litterfall, litter C and N content, fine root biomass, root C and N content, soil temperature, and soil moisture correlated with the soil MBC and MBN contents. All statistical analyses were performed using the PASW Statistics 18.0 for Windows.
3 Results
3.1 Environmental variables
The soil pH, soil C and N content, and soil C: N ratio under different forests all decreased with the increase of soil depth (
| Soil depth (cm) | Forest types | Soil pH | Bulk density (g cm-3) | Soil C content (g kg-1) | Soil N content (g kg-1) | Soil C: N |
|---|---|---|---|---|---|---|
| 0-10 | 5.00±0.08b | 1.36±0.04b | 6.65±2.67b | 0.54±0.09b | 11.97±2.91a | |
| 4.65±0.13c | 1.23±0.06cd | 4.33±1.21b | 0.53±0.04bc | 8.12±1.80b | ||
| 4.73±0.12c | 1.28±0.10bc | 4.08±0.35b | 0.53±0.05bc | 7.7±0.08b | ||
| 4.71±0.08c | 1.45±0.04a | 3.17±0.69b | 0.27±0.05c | 11.58±1.08a | ||
| Secondary forest | 6.57±0.22a | 1.17±0.04d | 12.71±4.94a | 1.26±0.34a | 9.91±1.23ab | |
| 10-20 | 4.71±0.18b | 1.47±0.04a | 1.85±0.34b | 0.31±0.07b | 6.16±0.91a | |
| 4.72±0.19b | 1.43±0.02a | 1.96±0.51b | 0.31±0.02b | 6.32±1.50a | ||
| 4.76±0.09b | 1.39±0.09a | 1.68±0.13b | 0.31±0.03b | 5.46±0.30a | ||
| 4.92±0.05b | 1.47±0.04a | 1.25±0.22b | 0.14±0.02c | 9.27±1.26a | ||
| Secondary forest | 6.50±0.69a | 1.31±0.02b | 4.57±0.93a | 0.54±0.09a | 8.44±1.15a |
Table 2.
Soil pH, soil bulk density, soil C and N content, and soil C: N ratio for the different forests (N=4)
In the subsoil, the soil pH and soil C and N concentrations in the secondary forest were all higher than those of the plantations, whereas the soil bulk density was lower than those of the plantations. No significant differences were found in soil pH, soil C content, or soil bulk density among the different plantations. In terms of soil N content, there was no significant difference among Eucalyptus, Pine, and Acacia plantations, which were each significantly higher than that of Casuarinas, and no significant difference was observed in the soil C: N ratio among the different forests.
The temperatures of topsoil and subsoil were both higher in summer and autumn and lower in spring and winter, which is consistent with the general rule (
![]()
Figure 3.Fig. 3
3.2 Seasonal variations of MBC and MBN
No significant difference was found in the seasonal variations of MBC content in the topsoil of Pine and Casuarinas plantations or in the subsoil of Acacia plantation, except that the MBC contents under different forests varied greatly among the different seasons (
The MBN content of different forests showed obvious seasonal variations, except for Eucalyptus (
![]()
Figure 4.Fig. 4
3.3 MBC and MBN under different forests
The annual mean MBC and MBN contents under different forests decreased with the increase of soil depth (
![]()
Figure 5.Fig. 5
The soil MBC: MBN ratio in the topsoil of Eucalyptus was significantly higher than that of the subsoil. However, no significant difference was observed between topsoil and subsoil in the other forests. In the topsoil, the soil MBC: MBN ratio of Eucalyptus was significantly higher than those of the other forests, and no significant difference was observed among other forests. In the subsoil, the corresponding ratio was highest in Pine, followed by Eucalyptus and Acacia plantations, and lowest in secondary forest and Casuarinas (
![]()
Figure 6.Fig. 6
The percentages of soil MBC and MBN within TC and TN, respectively, in topsoil were all lower than those in subsoil under each of the different forests. The percentages of soil MBC within TC under Eucalyptus, Pine, Casuarinas, and secondary forest differed significantly between topsoil and subsoil. The percentages of soil MBN within TN under Eucalyptus, Acacia and secondary forest differed significantly between topsoil and subsoil. Specifically, the percentage of soil MBC within TC in the subsoil of Eucalyptus was significantly higher than that of Casuarinas. In addition, no significant differences were found among different forests either in topsoil and subsoil. The percentage of soil MBN within TN in the topsoil of Casuarinas was significantly higher than those of Eucalyptus, Acacia and Pine, and the corresponding value in secondary forest was also higher than that of Pine. However, no significant differences were found among different forests either in topsoil and subsoil (
![]()
Figure 7.Fig. 7
3.4 Relationship between soil MBC and MBN contents and environmental factors
The effects of soil temperature and water content on the seasonal changes of soil MBC and MBN vary with forest types. However, the seasonal variations of soil MBC and MBN under different forests were all negatively correlated with soil temperature and positively correlated with soil water content (
| Forest types | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0-10 cm | 10-20 cm | 0-10 cm | 10-20 cm | 0-10 cm | 10-20 cm | 0-10 cm | 10-20 cm | |
| -0.216 | -0.007 | -0.421 | -0.937** | 0.388 | 0.170 | 0.346 | 0.760** | |
| -0.027 | -0.151 | -0.629** | -0.709** | 0.257 | 0.576* | 0.066 | 0.522* | |
| -0.640** | -0.217 | -0.857** | -0.786** | 0.537* | 0.057 | 0.553* | 0.082 | |
| -0.269 | -0.132 | -0.785** | -0.773** | 0.100 | 0.352 | 0.619* | 0.592* | |
| Secondary forest | -0.178 | -0.346 | -0.158 | -0.413 | 0.734** | 0.713** | 0.618* | 0.819** |
Table 3.
Pearson correlations between seasonal variations of soil MBC and MBN and soil temperature as well as water content
The MBC and MBN contents under different forests were significantly and positively correlated with TC, pH and litter N, and negatively correlated with soil bulk density and litter C. In addition, the MBN content was also significantly and positively correlated with root N both in topsoil and subsoil, and the MBC content in the subsoil was significantly and negatively correlated with root C (
| Soil depth (cm) | Variable | MBN | TC | TN | Bulk density | pH | Litterfall | Litter C | Litter N | Root biomass | Root C | Root N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0-10 | MBC | 0.750** | 0.697** | -0.030 | -0.609** | 0.899** | 0.202 | -0.500* | 0.651** | 0.301 | -0.388 | 0.326 |
| MBN | 1 | 0.820** | -0.050 | -0.586** | 0.915** | 0.408 | -0.624** | 0.671** | 0.151 | -0.188 | 0.505* | |
| 10-20 | MBC | 0.855** | 0.904** | 0.274 | -0.585** | 0.787** | 0.474* | -0.459* | 0.447* | 0.113 | -0.453* | 0.178 |
| MBN | 1 | 0.926** | -0.130 | -0.646** | 0.952** | 0.419 | -0.593** | 0.643** | 0.243 | -0.215 | 0.510* |
Table 4.
Pearson correlations between soil MBC and MBN and properties of soil, litter and fine roots
4 Discussion
4.1 Soil C and N contents under different forests
The qualities and quantities of soil C and soil N can be affected by forest types due to the variations in tree species composition, litter quantity and quality, root turnover rate, as well as root exudates in different forests (
4.2 MBC and MBN contents, and MBC/MBN ratios under different forests
The soil microbial biomass varies greatly among different forests, which is the combined result of tree species and environmental factors (
The mean MBC and MBN contents decreased with the increase of soil depth under the different forests, which is consistent with previous findings (
The MBC: MBN ratio reflects the relative ratio between fungi and bacteria in the soil (
The percentages of soil MBC and MBN within TC and TN in topsoil, respectively, were lower than those in subsoil under the same forests, which indicate that the carbon and nitrogen accumulations in the subsoil are higher than in the topsoil, and the soil organic matter is transferred from topsoil to subsoil (
4.3 Seasonal variation of MBC and MBN and its influencing factors
In this study, the MBC and MBN contents of different forests showed similar seasonal variations, which were single peak curves with higher values in spring or winter and lower values in summer or autumn. This agrees with the results from the coastal sandy dunes of Taiwan (
The soil pH is considered to be the main factor driving the spatial distribution of a soil microbial community, and pH is closely related to the microbial activity; when the pH value is lower than 7, the soil microbial activity increases gradually with an increase of soil pH (
5 Conclusions
The differences in soil microbial biomass C and N contents and their seasonal dynamics under five forest types (a secondary forest of L. glutinosa, and Casuarinas, Pine, Acacia, and Eucalyptus plantations) in the southeastern coast of Fujian province were investigated. The seasonal variations showed MBC and MBN contents which were all higher in spring and winter but lower in summer and autumn, and the seasonal variations of MBC and MBN were negatively correlated with soil temperature and positively correlated with soil water content, which indicated that the amount of microbial biomass in the sand dune forest ecosystem was largely controlled by soil moisture and soil temperature.
The mean MBC and MBN contents decreased with an increase of soil depth across the five forest types. Specifically, the MBC and MBN contents under secondary forest were significantly higher than those of plantations among the different soil layers; the MBC content under Casuarinas was the lowest in both topsoil and subsoil. These results imply that the long-term management of Casuarinas plantations led to soil decline, and the introduction of native tree species can modify soil parameters and consequently influence the belowground microbial community.
The MBC and MBN contents were significantly positively correlated with TC, pH, and litter N, and negatively correlated with soil bulk density and litter C. In addition, the MBN content was also significantly positively correlated with root N in both topsoil and subsoil, and the MBC content in the subsoil was significantly negatively correlated with root C. We found that several factors—environmental factors and the differences in litter and fine roots, soil nutrient contents, as well as the soil physical and chemical properties caused by different tree species—acted together to affect the concentrations of the soil MBC and MBN.
References
[4] L Bréchet, S Ponton, J Roy et al. Do tree species characteristics influence soil respiration in tropical forests? A test based on 16 tree species planted in monospecific plots. Plant & Soil, 319, 235-246(2009).
[5] C Brookes P, A Landman, G Pruden et al. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology & Biochemistry, 17, 837-842(1985).
[6] T Camenzind, S Httenschwiler, K Treseder K et al. Nutrient limitation of soil microbial processes in tropical forests. Ecological Monographs, 88, 4-21(2018).
[7] H Chang E, T Chen C, H Chen T et al. Soil microbial communities and activities in sand dunes of subtropical coastal forests. Applied Soil Ecology, 49, 256-262(2011).
[8] H Chen T, Y Chiu C, G Tian. Seasonal dynamics of soil microbial biomass in coastal sand dune forest. Pedobiologia, 49, 645-653(2005).
[9] M Chodak, M Pietrzykowski, K Sroka. Physiological profiles of microbial communities in mine soils afforested with different tree species. Ecological Engineering, 81, 462-470(2015).
[11] A Cregger M, W Schadt C, G Mcdowell N et al. Response of the soil microbial community to changes in precipitation in a semiarid ecosystem. Applied & Environmental Microbiology, 78, 8587-8594(2012).
[12] L Deng, P Shangguan Z. Afforestation drives soil carbon and nitrogen changes in China. Land Degradation & Development, 28, 151-165(2017).
[13] Y Feng, P Grogan, G Caporaso J et al. pH is a good predictor of the distribution of anoxygenic purple phototrophic bacteria in Arctic soils. Soil Biology & Biochemistry, 74, 193-200(2014).
[15] W Gao, Q Huang Z, F Ye G et al. Effects of forest cover types and environmental factors on soil respiration dynamics in a coastal sand dune of subtropical China. Journal of Forestry Research, 29, 1645-1655(2018).
[16] F Harris R. Effect of water potential on microbial growth and activity. In: Parr J F, Gardner W R, Elliott L F (eds.). Water potential relations in soil microbiology, Special publication no 9, 23-96(1980).
[17] M Hoogmoed, C Cunningham S, J Baker P et al. Is there more soil carbon under nitrogen-fixing trees than under non-nitrogen-fixing trees in mixed-species restoration plantings?. Agriculture Ecosystems & Environment, 188, 80-84(2014).
[18] S Hu, C Coleman D, R Carroll C et al. Labile soil carbon pools in subtropical forest and agricultural ecosystems as influenced by management practices and vegetation types. Agriculture Ecosystems & Environment, 65, 69-78(1997).
[19] R Jandl, M Lindner, L Vesterdal et al. How strongly can forest management influence soil carbon sequestration?. Geoderma, 137, 253-268(2007).
[20] G Joergensen R, T Müller. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the
[21] G Joergensen R, T Müller. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the
[22] L Karlen D, J Mausbach M, W Doran J et al. Soil quality: A concept, definition, and framework for evaluation. Soil Science Society of America Journal, 61, 4-10(1997).
[23] J Landesman W, M Nelson D, C Fitzpatrick M. Soil properties and tree species drive ß-diversity of soil bacterial communities. Soil Biology & Biochemistry, 76, 201-209(2014).
[24] L Li S, X Fang, H Xiang W et al. Soil microbial biomass carbon and nitrogen concentrations in four subtropical forests in hilly region of central Hunan Province, China. Scientia Silvae Sinicae, 50, 8-16(2014).
[25] G Liu C, Q Jin Y, N Hu Y et al. Drivers of soil bacterial community structure and diversity in tropical agroforestry systems. Agriculture Ecosystems & Environment, 278, 24-34(2019).
[26] S Liu, K Wang C. Spatio-temporal patterns of soil microbial biomass carbon and nitrogen in five temperate forest ecosystems. Acta Ecologica Sinica, 30, 3135-3143(2010).
[27] W Luan J, R Liu S, X Wang J et al. Rhizospheric and heterotrophic respiration of a warm-temperate oak chronosequence in China. Soil Biology & Biochemistry, 43, 503-512(2011).
[28] S Manzoni, P Schimel J, A Porporato. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology, 93, 930-938(2012).
[31] P Nannipieri, J Ascher, T Ceccherini M et al. Microbial diversity and soil functions. European Journal of Soil Science,, 54, 655-670(2003).
[32] C Pérez-Cruzado, P Mansilla-Salinero, R Rodríguez-Soalleiro et al. Influence of tree species on carbon sequestration in afforested pastures in a humid temperate region. Plant & Soil, 353, 333-353(2012).
[33] S Parker S, P Schimel J. Soil nitrogen availability and transformations differ between the summer and the growing season in a California grassland. Applied Soil Ecology, 48, 185-192(2011).
[34] I Paul K, J Polglase P, G Nyakuengama J et al. Change in soil carbon following afforestation. Forest Ecology & Management, 168, 241-257(2002).
[35] L Pearson H, M Vitousek P. Stand dynamics, nitrogen accumulation, and symbiotic nitrogen fixation in regenerating stands of
[36] J Qiu S, T Ju X, J Ingwersen et al. Changes in soil carbon and nitrogen pools after shifting from conventional cereal to greenhouse vegetable production. Soil & Tillage Research, 107, 80-87(2010).
[37] C Resh S, D Binkley, A Parrotta J. Greater soil carbon sequestration under nitrogen fixing trees compared with
[38] U Sarathchandra S, W Perrott K, R Boase M et al. Seasonal changes and the effects of fertiliser on some chemical, biochemical and microbiological characteristics of high-producing pastoral soil. Biology & Fertility of Soils, 6, 328-335(1988).
[39] C Shen C, B Xiong J, Y Zhang H et al. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biology & Biochemistry, 57, 204-211(2013).
[40] Y Steinberger, L Zelles, Y Bai Q et al. Phospholipid fatty acid profiles as indicators for the microbial community structure in soils along a climatic transect in the Judean Desert. Biology & Fertility of Soils, 28, 292-300(1999).
[41] F Tam P C. Mycorrhizal associations in
[42] H Templer P, E Dawson T. Nitrogen uptake by four tree species of the Catskill Mountains, New York: Implications for forest N dynamics. Plant & Soil, 262, 251-261(2004).
[43] N Ussiri D A, R Lal, A Jacinthe P. Soil properties and carbon sequestration of afforested pastures in reclaimed mine soils of Ohio. Soil Science Society of America Journal, 70, 1797-1806(2006).
[44] M Wang F, A Li Z, P Xia H et al. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Science & Plant Nutrition, 56, 297-306(2010).
[45] H Wang, R Liu S, X Wang J et al. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. Forest Ecology & Management, 300, 4-13(2013).
[46] H Wang M, H Wan X, P Yu Z et al. a. Effects of tree species transition on soil microbial biomass and community structure in subtropical China. Acta Ecologica Sinica, 36, 417-423(2016).
[47] N Wang, X Yang, L Li S et al. b. Seasonal dynamics of soil microbial biomass carbon-nitrogen in the Korean Pine mixed forests along elevation gradient. Scientia Silvae Sinicae, 52, 150-158(2016).
[48] A Wardle D. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil. Biological Reviews, 67, 321-358(2010).
[49] S Xiao S, F Ye G, H Zhang L et al. Soil heterotrophic respiration in
[50] Q Xu H, E Zhang J, J Feng L et al. Effects of different land use patterns on microbial biomass carbon and nitrogen in Guangdong Province. Acta Ecologica Sinica, 29, 4112-4118(2009).
[51] K Yang, J Zhu J, X Zhang J et al. Seasonal dynamics of soil microbial biomass C and N in two larch plantation forests with different ages in Northeastern China. Acta Ecologica Sinica, 29, 5500-5507(2009).
[52] S Yang Y, S Chen G, P Lin et al. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in subtropical China. Annals of Forest Science, 61, 617-627(2004).
[53] F Ye G, J Zhang S, H Zhang L et al. Age-related changes in nutrient resorption patterns and tannin concentration of
[57] H Zhu T, J Zhang, X Hu T et al. Mycorrhizae type associated with Eucalyptus in Sichuan and effect of forest density on mycorrhizal fungi. Journal of Sichuan Agricultural University, 19, 222-224(2001).

Set citation alerts for the article
Please enter your email address
© Copyright 2018-2021 | Chinese Laser Press. All Rights Reserved 沪ICP备15018463号-20